I.2 Фотосинтез, необходимые для него условия. Темновая фаза фотосинтеза Монооксигеназные реакции необходимы для
Для темновых реакций, которые протекают в строме, свет не нужен. Восстановление СО 2 происходит за счет энергии (АТФ) и восстановительной силы (НАДФ·Н 2), образующихся при световых реакциях. Темновые реакции контролируются ферментами. Последовательность этих реакций была определена в США Кальвином, Бенсоном и Бэссемом в период с 1946 по 1953 г.; в 1961 г. Кальвину была присуждена за эту работу Нобелевская премия.
Эксперименты Кальвина
Работы Кальвина были основаны на использовании радиоактивного изотопа углерода 14 С (период полураспада 5570 лет, см. Приложение 1.3), который стал доступным для исследователей только в 1945 г. Кроме того, Кальвин применял бумажную хроматографию, которая в то время была сравнительно новым, еще мало распространенным методом. Культуры одноклеточной зеленой водоросли хлореллы (Chlorella) выращивали в специальном аппарате (рис. 9.17). Культуру выдерживали с 14 СО 2 в течение различных промежутков времени, затем клетки быстро фиксировали, выливая суспензию в горячий метанол. Растворимые продукты фотосинтеза экстрагировали, концентрировали и разделяли при помощи двумерной хроматографии на бумаге (рис. 9.18 и Приложение 1.8.2). Цель состояла в том, чтобы проследить путь, по которому меченый углерод попадает (через ряд промежуточных продуктов) в конечные продукты фотосинтеза. Положение радиоактивных соединений на бумаге определяли с помощью радиоавтографии : для этого на хроматограмму накладывали фотопленку, чувствительную к излучению 14 С, и она засвечивалась, т. е. чернела, в тех местах, где находились радиоактивные вещества (рис. 9.18). Уже за одну минуту инкубации с 14 СО 2 синтезировались многие сахара и органические кислоты, в том числе различные аминокислоты. Однако Кальвину удалось, используя очень короткие экспозиции - в течение 5 и менее секунд - идентифицировать первый продукт фотосинтеза и установить, что это кислота, содержащая три атома углерода, а именно фосфоглицериновая кислота (ФГК). Затем он выяснил всю цепь промежуточных соединений, по которой передается фиксированный углерод; эти стадии будут рассмотрены позднее. С тех пор эти реакции называют циклом Кальвина (или циклом Кальвина - Бенсона- Бэссема).

Рис. 9.18. А. Фиксация 14 СО 2 У водорослей при кратковременном освещении. Определение продуктов фиксации с помощью хроматографии на бумаге и радиоавтографии. Б. Радиоавтографы продуктов фотосинтеза, получающихся после кратковременного освещения водорослей в присутствии 14 СО 2
9.18. Какие преимущества дает применение долгоживущих радиоактивных изотопов в биологических исследованиях?
9.19. Какие преимущества можно получить, взяв хлореллу вместо высшего растения?
9.20. Почему сосуд аппарата Кальвина имеет плоскую, а не шарообразную форму?
Этапы пути углерода
Фиксация двуокиси углерода:

Акцептором СО 2 служит пятиуглеродный сахар (пентоза) рибулозобисфосфат (т. е. рибулоза с двумя фосфатными группами; раньше это соединение называли рибулозодифосфатом). Присоединение СО 2 к тому или иному веществу называется карбоксилированием , а фермент, катализирующий такую реакцию, - карбоксилазой . Образующийся шестиуглеродный продукт неустойчив и сразу же распадается на две молекулы фосфоглицериновой кислоты (ФГК), которая и является первым продуктом фотосинтеза. Фермент рибулозобисфосфат-карбоксилаза содержится в строме хлоропластов в большом количестве - это фактически самый распространенный в мире белок.
Восстановительная фаза :

ФГК содержит три атома углерода и имеет кислотную карбоксильную группу (-СООН). ТФ - это триозофосфат, или глицеральдегидфосфат (трехуглеродный сахар); он имеет альдегидную группу (-СНО).
Для удаления кислорода из ФГК (т. е. для ее восстановления) используются восстановительная сила НАДФ·Н 2 и энергия АТФ. Реакция протекает в два этапа: сначала расходуется часть АТФ, образовавшегося в ходе световых реакций, а затем используется весь НАДФ·Н 2 , также полученный на свету. Суммарный результат - восстановление карбоксильной группы кислоты (-СООН) до альдегидной группы (-СНО). Продукт реакции - триозофосфат, т. е. трехуглеродный сахар с присоединенной к нему фосфатной группой. В этом соединении больше химической энергии, чем в ФГК, и это первый углевод, который образуется при фотосинтезе.
Регенерация акцептора для СО 2 - рибулозобисфосфата . Часть триозофосфата (ТФ) должна израсходоваться на регенерацию рибулозобисфосфата, который используется в первой реакции. Этот процесс представляет собой сложный цикл, в котором участвуют сахарофосфаты с 3, 4, 5, 6, 7 атомами углерода. Именно здесь и расходуется остальной АТФ. Все темновые реакции суммированы на рис. 9.19. На этом рисунке цикл Кальвина изображен в виде "черного ящика", в который с одной стороны поступают СО 2 и Н 2 О, а с другой стороны выходит триозофосфат. Как видно из этой схемы, остаток АТФ используется для фосфорилирования рибулозобисфосфата, однако детали этой сложной цепи реакций не показаны.

Из рис. 9.19 можно вывести такое суммарное уравнение:

Здесь важно обратить внимание на то, что на образование двух молекул триозофосфата идет шесть молекул СО 2 . Уравнение можно упростить, поделив все коэффициенты на 6:

9.21. Перерисуйте рис. 9.19, указав только число атомов углерода, участвующих в реакциях; например, вместо 6 РиБФ напишите "6 × 5С" и т. д.
Основные сведения о процессе фотосинтеза резюмированы в табл. 9.6.
| Световые реакции | Темновые реакции | |
| Локализация в хлоропластах | Тилакоиды | Строма |
| Реакции | Фотохимические, т. е. требуют света. Световая энергия вызывает перенос электронов от "доноров" электронов к их "акцепторам" либо по нециклическому, либо по циклическому пути. Участвуют две фотосистемы - Ι и ΙΙ. В них находятся молекулы хлорофиллов, которые при поглощении энергии света испускают электроны. Вода служит донором электронов для нециклического пути. Перенос электронов приводит к образованию АТФ (фотофосфорилированию) и НАДФ·Н 2 (см. также табл. 9.5). | Не требуют света. СО 2 фиксируется, когда связывается с пятиуглеродным акцептором - рибулозобисфосфатом (РиБФ); при этом образуются две молекулы трехуглеродного соединения-фосфоглицериновой кислоты (ФГК), первого продукта фотосинтеза. Происходит целый ряд реакций, в совокупности называемых циклом Кальвина; при этом регенерируется акцептор для СО 2 -РиБФ, а ФГК восстанавливается, превращаясь в сахар (см. также рис. 9.19). |
| Комбинированные уравнения |
Монооксигеназные реакции необходимы для:
1. специфических превращений аминокислот, например, для синтеза тирозина из фенилаланина (фермент – фенилаланингидроксилаза);
2. синтеза холестерола, желчных кислот в печени; стероидных гормонов в коре надпочечников, яичниках, плаценте, семенниках; витамина D 3 в почках;
3. обезвреживания чужеродных веществ (ксенобиотиков) в печени.
Ферменты монооксигеназного пути окисления локализованы в мембранах эндоплазматического ретикулума (при гомогенизации тканей эти мембраны превращаются в микросомы – мембранные пузырьки). Поэтому монооксигеназный путь окисления называют микросомальным окислением.
Микросомальное окисление представляет короткую электронтранспортную цепь, включающую НАДФ, ФАД, ФМН, цитохром Р 450 .
Микросомальная система включает два фермента: цитохром Р 450 и НАДФН-цитохром-Р 450 -редуктазу.
НАДФН-цитохром Р 450 – редуктаза – флавопротеин, в качестве простетической группы содержит два кофермента ФАД и ФМН.
Цитохром Р 450 – гемопротеин, содержит простетическую группу гем и участки связывания для кислорода и субстрата. Восстановленный цитохром Р 450 имеет максимум поглощения при 450 нм. Выполняет две функции: связывание окисляемого субстрата и активация молекулярного кислорода.

Рис. 11.1. Схема микросомального окисления
Микросомальное окисление протекает в несколько этапов:
1. связывание в активном центре цитохрома Р 450 субстрата RН;
2. присоединение первого электрона и восстановление железа в геме до Fe 2+ ; изменение валентности железа увеличивает сродство комплекса Р 450 – Fe 2+ RH к молекуле кислорода; присоединение второго электрона к молекуле кислорода и образование неустойчивого пероксикомплекса Р 450 –Fe 2+ О 2 - RH;
3. Fe 2+ окисляется, при этом электрон присоединяется к молекуле кислорода; восстановленный атом кислорода (О 2 -) связывает два протона (донор протонов – НАДФН+Н +) и образуется 1 молекула воды; второй атом кислорода участвует в гидроксилировании субстрата RH; гидроксилированный субстрат ROH отделяется от фермента.
В результате гидроксилирования гидрофобный субстрат становится более полярным, повышается его растворимость и возможность выведения из организма с мочой. Так окисляются многие ксенобиотики, лекарственные вещества.
В редких случаях в результате гидроксилирования токсичность соединения увеличивается. Например, при окислении нетоксичного бензпирена (содержится в табачном дыму, копченостях) образуется токсичный оксибензпирен, который является сильным концерогеном, индуцирующим злокачественное перерождение клеток.
В митохондриях содержится монооксигеназная система, которая выполняет биосинтетическую функцию: синтез холестерола; стероидных гормонов (кора надпочечников, яичники, плацента, семенники); желчных кислот (печень); образование витамина D 3 (почки).
Рис. 12. Схема переноса электронов хлорофилла в процессе синтеза АТФ (фосфорилирования). При нециклическом фосфорилировании за счет энергии ФС II идет синтез АТФ, а за счет ФС I идет синтез НАДФ · 2Н. При циклическом фосфорилировании работает только ФС I и происходит синтез АТФ
Под действием света происходит возбуждение молекул хлорофилла ФС I и ФС II, которые, поглощая кванты света, испускают возбужденные электроны. За счет энергии электронов, летящих по цепи ферментов от ФС II к ФС I, происходит синтез молекул АТФ. Этот процесс называется фосфорилированием и происходит в присутствии фермента АТФ-синтетазы.
АДФ + Фн АТФ
где Фн - неорганический фосфат H3 PO4 .
Электроны ФС II, потерявшие энергию, попадают на ФС I, которая является их конечным акцептором.
Электроны, покидающие хлорофилл фотосистемы ФС I при поглощении света, также попадают на электроннотранспортную цепь. Но за счет их энергии идет соединение протонов Н+ с переносчиком НАДФ+ .
Электроны, которые теряет ФС I, восполняются электронами ФС II, а электроны, отданные молекулой воды при фотолизе, восполняют электроны, потерянные хлорофиллом ФС II, и он также восстанавливается:
хлорофилл+2 + 2ē хлорофилл.
На мембране имеются специальные протонные каналы, по которым в определенный момент ионы водорода могут переходить из Н+ -резервуара тилакоида в строму хлоропласта. Каналы связаны с ферментом АТФ-синтетазой. Когда возбужденные электроны ФС I, двигаясь по мембране, достигают протонного канала, он открывается и в него устремляются ионы водорода. Этот процесс сопряжен с синтезом АТФ и происходит синхронно.
С наружной стороны мембраны тилакоида, т. е. в строме хлоропласта, скапливаются молекулы переносчика водорода НАДФ+ в окисленном состоянии. Они принимают электроны от ФС I, за счет чего происходит их соединение с ионами водорода Н+ и образование НАДФ · 2H:
НАДФ+ + 2H+ + 2ē НАДФ · 2Н. (4)
Синтез АТФ и НАДФ · 2Н протекает на мембранах тилакоидов и сопряжен с переносом возбужденных электронов по электронно-транспортной цепи. Таким образом, энергия солнца преобразуется в энергию возбужденных электронов, а далее запасается в процессе синтеза в молекулах АТФ и НАДФ · 2Н.
Суммарное уравнение реакций световой фазы:
H2 O + НАДФ+ + 2АДФ + 2Фн НАДФ · 2Н + 2АТФ + 1 /2 O2 .
Темновая фаза
Реакции темновой фазы (рис. 13) протекают в строме хлоропластов, куда поступают молекулы НАДФ · 2Н и АТФ, синтезированные в световую фазу, и углекислый газ из атмосферы. Последовательность циклических реакций этой фазы была описана американским ученым-биохимиком Мэлвином Кальвином и получила название цикла Кальвина. Здесь происходит связывание молекул CO2 , активирование соединений за счет АТФ (фосфорилирование), восстановление углерода водородом из НАДФ · 2Н и синтез глюкозы. Источником энергии являются синтезированные на первой стадии молекулы АТФ.
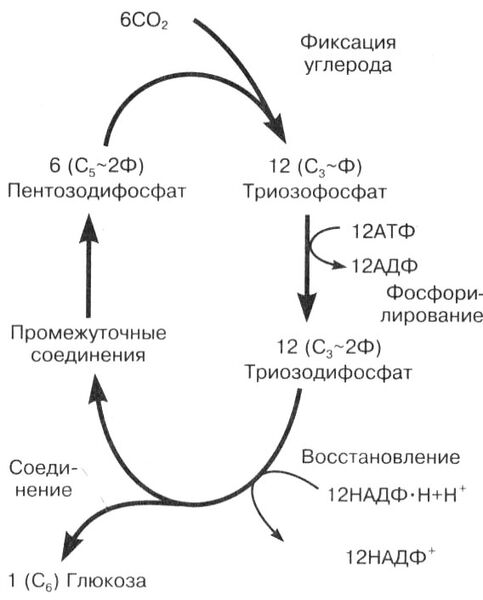
Рис. 13. Общая схема темновых реакций фотосинтеза. Цикл Кальвина
В строме хлоропласта постоянно присутствует пятиуглеродный углевод (пентоза), связанный с двумя остатками фосфорной кислоты - рибулозодифосфат.
Образующееся шестиуглеродное соединение неустойчиво и сразу же распадается на два триозофосфата.
С5 -углевод-2Ф + CO2 С6 -углевод-2Ф 2С3 -Ф
2С3 Ф + 2АТФ 2АДФ + 2С3 ~2Ф.

Рис. 14. Фиксация углерода, его фосфорилирование и восстановление
После этого происходит восстановление триозодифосфатов молекулами НАДФ · 2Н:
2С3 ~2Ф + 2НАДФ · 2Н 2С3 + 2НАДФ+ + 2Ф.
Две молекулы триозы соединяются между собой, и образуется глюкоза, которая может в дальнейшем превращаться в сахарозу, крахмал и другие полисахариды:
2С3 С6 Н12 O6 .
Часть молекул триоз может использоваться для синтеза аминокислот, глицерина, высших жирных кислот.
Частично триозы продолжают участвовать в циклических реакциях и превращаются вновь в пентозу, которая замыкает цикл.
В реакции участвуют одновременно шесть молекул каждого вещества. Таким образом, для синтеза одной полностью новой молекулы глюкозы цикл должен повториться шесть раз, т. е. должно усвоиться шесть молекул CO2 .
Освобожденные молекулы АТФ и НАДФ+ вновь возвращаются к мембранам тилакоидов для участия в световых реакциях.
Влияние на скорость фотосинтеза различных факторов
Интенсивность фотосинтеза зависит от целого ряда факторов. Во-первых, это длина световой волны.
Наиболее интенсивен процесс в ультрафиолетовой и красной части спектра. Кроме того, скорость фотосинтеза зависит от степени освещенности, и до определенной величины она возрастает пропорционально освещенности, но далее уже не зависит от нее (рис. 15).

Рис. 15. Влияние различных факторов на скорость фотосинтеза
Другим фактором является концентрация углекислого газа. Чем выше количество углекислого газа, тем интенсивнее идет процесс фотосинтеза. В обычных условиях недостаток углекислого газа является главным лимитирующим фактором, так как в атмосферном воздухе содержится небольшой его процент. Однако в условиях теплиц можно устранить этот дефицит, что благоприятно влияет на интенсивность роста и развития растений.
Немаловажным фактором является температура.
Все реакции фотосинтеза катализируются ферментами, для которых оптимальной температурой является интервал 25–30 °C. При низких температурах скорость действия ферментов резко снижается.
Вода является также важным фактором, влияющим на фотосинтез. Однако оценить количественно этот фактор невозможно, так как она участвует во многих других обменных процессах.
I.2 Фотосинтез, необходимые для него условия
Фотосинтез у зеленых растений – это процесс преобразования света в химическую энергию органических соединений, синтезируемых из диоксида углерода и воды. Процесс фотосинтеза представляет собой цепь окислительно-восстановительных реакций, совокупность которых подразделяют на две фазы – световую и темновую.
1. Световая фаза. Для этой фазы характерно то, что энергия солнечной радиации, поглощенная пигментами системы хлоропластов, преобразуется в электрохимическую.
При действии света на хлоропласт начинается электронный поток по системе переносчиков – сложных органических соединений, встроенных в мембраны тилакоидов. С переносом электронов по ЭТЦ сопряжено активное поступление протонов через тилакоидную мембрану из стромы внутрь тилакоида. В тилакоидном пространстве происходит увеличение концентрации протонов за счет расщепления молекул воды и в результате окисления электронного переносчика пластохинона на внутренней стороне мембраны. Когда протоны идут обратно по градиенту из тилакоидного пространства в строму, на наружной поверхности тилакоида с участием фермента АТФ-синтетазы из АДФ и фосфорной кислоты синтезируется АТФ, т. е. происходит фотосинтетическое фосфореилирование с запасанием энергии в АТФ, которая затем переходит в строму хлоропласта.
Заканчивается передача электронов следующим образом. Достигнув внешней поверхности мембраны тилакоида, пара электронов следует с ионом водорода, находящимся в строме. Оба электрона и ион водорода присоеденяются к молекуле переносчика водорода – НАДФ+ (никатиномидадениндинуклетидфосфат), который при этом переходит в свою востановленную форму
НАДФ Н+Н+:
НАДФ++2Н++2е-→НАДФ Н+Н+
Следовательно активированные световой энергией электроны используются на присоедининие атома водорода к его переносчику, т. е. на восстановление НАДФ+ в НАДФ Н+Н+, который с наружной поверхности фотосинтетической мембраны переходит в строму.
В молекулах хлорофилла, утративших свои электроны, образовавшиеся электронные «дырки» действуют как сильный окислитель и отрывают электроны от молекул воды. Через ряд переносчиков эти электроны передаются на молекулу хлорофилла и заполняют «дырку». Внутри тилакоида происходит фотоокислние (фотолиз) воды, в результате которого выделяется свободный кислород, а также накапливаются ионы водорода
2Н2О→4Н++4е-+О2
Таким образом, во время световой фазы фотосинтеза происходят три процесса: образование кислорода вследствие разложения воды, синтез АТФ и образование атомов водорода в форме НАДФ Н2. Кислород диффундирует в атмосферу, а АТФ и НАДФ Н2 транспортируются в матрикс пластид и участвуют в процессе темновой фазы.
2.Темновая фаза фотосинтеза протекает в матриксе хлоропласта как на свету, так и в темноте и представляет собой ряд последовательных преобразований СО2, поступаещего из воздуха. Осуществляются реакции темновой фазы за счет энергии АТФ и НАДФ Н2 и использовании имеющихся в пластидах пятиуглеродных сахаров, один из которых – рибулозодифосфат – является акцептором СО2. Ферменты связывают пятиуглеродный сахар с углекислым газом воздуха. При этом образуются соединения которые последовательно восстанавливаются до шестиуглеродной молекулы глюкозы.
Суммарная реакция фотосинтеза
6СО2+6Н2 энергия света С6Н12О6+6О2
Хлорофилл
В процессе фотосинтеза кроме моносахаридов (глюкоза и др.), которые превращаются в крахмал и запасаются растением, синтезируются мономеры других органических соединений – аминокислоты, глицерин и жирные кислоты. Таким образом, благодоря фотосинтезу растительные, а точнее – хлорофиллсодержащие, клетки обеспечивают себя и все живое на Земле необходимыми органическими веществами и кислородом.
I.3 Деление клетки
Описано три способа деления эукариотических клеток: амитоз (прямое деление), митоз (непрямое деление) и мейоз (редукционное деление).
Амитоз – относительно редкий способ деления клетки. При амитозе интерфазное ядро делится путем перетяжки, равномерное распределение наследственного материала не обеспечивается. Нередко ядро делится без последующего разделения цитоплазмы и образуются двухъядерные клетки. Клетка, претерпевшая амитоз, в дальнейшем не способна вступать в нормальный митотический цикл. Поэтому амитоз встречается, как правило, в клетках и тканях, обреченных на гибель.
Митоз. Митоз, или непрямое деление, - основной способ деления эукариотических клеток. Митоз – это деление ядра, которое приводит к образованию двух дочерних ядер, в каждом из которых имеется точно такой же набор хромосом, что и был в родительском ядре.
В непрерывном процессе митотического деления различают четыре фазы: профазу, метафазу, анафазу и телофазу.
Профаза – самая длительная фаза митоза, когда происходит перестройка всей структуры ядра для деления. В профазе происходит укорочение и утолщение хромосом вследствие их спирализации. В это время хромосомы двойные (удвоение происходит в S-периоде интерфазы), состоят из двух хроматид, связанных между собой в области первичной перетяжки осбой структурой – цетромерой. Одновременно с утолщением хромосом исчезает ядрышко и фрагментируется (распадается на отдельные цистерны) ядерная оболочка. После распада ядерной оболочки хромосомы свободно и беспорядочно лежат в цитоплазме. Начинается формирование ахромативного веретена – веретена деления, которое представляет систему нитей, идущих от полюсов клетки. Нити веретена имеют диаметр около 25нм. Это пучки микротрубочек, состоящих из субъедениц белка тубулина. Микротрубочки начинают формироваться со стороны центриолей либо со стороны хромосом (в клетках растений).
Метафаза. В метафазе завершается образование веретена деления, которое состоит из микротрубочек двух типов: хромосомных, которые связываются с центромерами хромосом, и ценросомных (полюсных), которые тянутся от полюса к полюсу клетки. Каждая двойная хромосома прикрепляется к микротрубочкам веретена деления. Хромосомы как бы выталкиваются микротрубочками в область экватора клетки, т.е. располагаются на равном расстоянии от полюсов. Они лежат в одной плоскости и образуют так называемую экваториальную, или метафазную пластинку. В метафазе отчетливо видно двойное строение хромосом, соединенных только в области центромеры. Именно в этот период легко подсчитать число хромосом, изучать их морфологические особенности.
Анафаза начинается делением центромеры. Каждая из хроматид одной хромосомы становится самостоятельной хромосомой. Сокращение тянущих нитей ахроматинового веретена увлекает их к противоположным полюсам клетки. В результате у каждого из полюсов клетки оказывается столько же хромосом, сколько было их в материнской клетке, причем набор их одинаков.
Телофаза – последняя фаза митоза. Хромосомы деспирализуются, становятся плохо заметными. На каждом из полюсов вокруг хромосом воссоздается ядерная оболочка. Формируются ядрышки, веретено деления исчезает. В образовавшихся ядрах каждая хромосома состоит теперь всего из одной хроматиды, а не из двух.
С использованием световой энергии или без нее. Он характерен для растений. Рассмотрим далее, что собой представляют темновая и световая фаза фотосинтеза.
Общие сведения
Органом фотосинтеза у высших растений является лист. В качестве органоидов выступают хлоропласты. В мембранах их тилакоидов присутствуют фотосинтетические пигменты. Ими являются каротиноиды и хлорофиллы. Последние существуют в нескольких видах (а, с, b, d). Главным из них считается а-хлорофилл. В его молекуле выделяется порфириновая "головка" с атомом магния, расположенным в центре, а также фитольный "хвост". Первый элемент представлен в виде плоской структуры. "Головка" является гидрофильной, поэтому располагается на той части мембраны, которая направлена к водной среде. Фитольный "хвост" является гидрофобным. За счет этого он удерживает хлорофилльную молекулу в мембране. Хлорофиллами поглощается сине-фиолетовый и красный свет. Они также отражают зеленый, за счет чего растения имеют характерный для них цвет. В мембранах тилактоидов молекулы хлорофилла организованы в фотосистемы. Для синезеленых водорослей и растений характерны системы 1 и 2. Фотосинтезирующие бактерии имеют только первую. Вторая система может разлагать Н 2 О, выделять кислород.

Световая фаза фотосинтеза
Процессы, происходящие в растениях, отличаются сложностью и многоступенчатостью. В частности, выделяют две группы реакций. Ими являются темновая и световая фаза фотосинтеза. Последняя протекает при участии фермента АТФ, белков, переносящих электроны, и хлорофилла. Световая фаза фотосинтеза происходит в мембранах тилактоидов. Хлорофилльные электроны возбуждаются и покидают молекулу. После этого они попадают на внешнюю поверхность мембраны тилактоида. Она, в свою очередь, заряжается отрицательно. После окисления начинается восстановление молекул хлорофилла. Они отбирают электроны у воды, которая присутствует во внутрилакоидном пространстве. Таким образом, световая фаза фотосинтеза протекает в мембране при распаде (фотолизе): Н 2 О + Q света → Н + + ОН —
Ионы гидроксила превращаются в реакционноспособные радикалы, отдавая свои электроны:
ОН — → .ОН + е —
ОН-радикалы объединяются и образуют свободный кислород и воду:
4НО. → 2Н 2 О + О 2 .

При этом кислород удаляется в окружающую (внешнюю) среду, а внутри тилактоида идет накопление протонов в особом "резервуаре". В результате там, где протекает световая фаза фотосинтеза, мембрана тилактоида за счет Н + с одной стороны получает положительный заряд. Вместе с этим за счет электронов она заряжается отрицательно.
Фосфирилирование АДФ
Там, где протекает световая фаза фотосинтеза, присутствует разность потенциалов между внутренней и наружной поверхностями мембраны. Когда она достигает 200 мВ, начинается проталкивание протонов сквозь каналы АТФ-синтетазы. Таким образом, световая фаза фотосинтеза происходит в мембране при фосфорилировании АДФ до АТФ. При этом атомарный водород направляется на восстановление особого переносчика никотинамидадениндинуклеотидфосфата НАДФ+ до НАДФ.Н2:
2Н + + 2е — + НАДФ → НАДФ.Н 2
Световая фаза фотосинтеза, таким образом, включает в себя фотолиз воды. Его, в свою очередь, сопровождают три важнейших реакции:
- Синтез АТФ.
- Образование НАДФ.Н 2 .
- Формирование кислорода.
Световая фаза фотосинтеза сопровождается выделением последнего в атмосферу. НАДФ.Н2 и АТФ перемещаются в строму хлоропласта. На этом световая фаза фотосинтеза завершается.

Другая группа реакций
Для темновой фазы фотосинтеза не нужна световая энергия. Она идет в строме хлоропласта. Реакции представлены в виде цепочки последовательно происходящих преобразований поступающего из воздуха углекислого газа. В итоге образуются глюкоза и прочие органические вещества. Первой реакцией является фиксация. В качестве акцептора углекислого газа выступает рибулозобифосфат (пятиуглеродный сахар) РиБФ. Катализатором в реакции является рибулозобифосфат-карбоксилаза (фермент). В результате карбоксилирования РиБФ формируется шестиуглеродное неустойчивое соединение. Оно практически мгновенно распадается на две молекулы ФГК (фосфоглицериновой кислоты). После этого идет цикл реакций, где она через несколько промежуточных продуктов трансформируется в глюкозу. В них используются энергии НАДФ.Н 2 и АТФ, которые были преобразованы, когда шла световая фаза фотосинтеза. Цикл указанных реакций именуется "циклом Кальвина". Его можно представить следующим образом:
6СО 2 + 24Н+ + АТФ → С 6 Н 12 О 6 + 6Н 2 О
Помимо глюкозы, в ходе фотосинтеза образуются прочие мономеры органических (сложных) соединений. К ним, в частности, относят жирные кислоты, глицерин, аминокислоты нуклеотиды.

С3-реакции
Они представляют собой тип фотосинтеза, при котором в качестве первого продукта образуются трехуглеродные соединения. Именно он описан выше как цикл Кальвина. В качестве характерных особенностей С3-фотосинтеза выступают:
- РиБФ является акцептором для углекислого газа.
- Реакция карбоксилирования катализирует РиБФ-карбоксилаза.
- Образуется шестиуглеродное вещество, которое впоследствии распадается на 2 ФГК.
Фосфоглицериновая кислота восстанавливается до ТФ (триозофосфатов). Часть из них направляется на регенерацию рибулозобифосфата, а остальная - превращается в глюкозу.
С4-реакции
Для этого типа фотосинтеза характерно появление четырехуглеродных соединений в качестве первого продукта. В 1965 году было выявлено, что С4-вещества появляются первыми у некоторых растений. Например, это было установлено для проса, сорго, сахарного тростника, кукурузы. Эти культуры стали именовать С4-растениями. В следующем, 1966-м, Слэк и Хэтч (австралийские ученые) выявили, что у них почти полностью отсутствует фотодыхание. Также было установлено, что такие С4 растения намного эффективнее осуществляют поглощение углекислого газа. В результате путь трансформации углерода в таких культурах стали именовать путем Хэтча-Слэка.

Заключение
Значение фотосинтеза очень велико. Благодаря ему из атмосферы ежегодно поглощается углекислый газ в огромных объемах (миллиардами тонн). Вместо него выделяется не меньшее количество кислорода. Фотосинтез выступает в качестве основного источника формирования органических соединений. Кислород участвует в образовании озонового слоя, обеспечивающего защиту живых организмов от воздействия коротковолновой УФ-радиации. В процессе фотосинтеза лист поглощает только 1% всей энергии света, падающего на него. Его продуктивность находится в пределах 1 г органического соединения на 1 кв. м поверхности за час.







